NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Walker HK, Hall WD, Hurst JW, editors. Clinical Methods: The History, Physical, and Laboratory Examinations. 3rd edition. Boston: Butterworths; 1990. Bookshelf ID: NBK365 PMID: 21250206
Definition
Normal ventilation is an automatic, seemingly effortless inspiratory expansion and expiratory contraction of the chest cage. This act of normal breathing has a relatively constant rate and inspiratory volume that together constitute normal respiratory rhythm. The accessory muscles of inspiration (sternocleidomastoid and scalenes) and expiration (abdominal) are not normally used in the resting state. Abnormalities may occur in rate, rhythm, and in the effort of breathing.Technique
The establishment of the tidal volume and pattern of respiration in normal individuals is a complicated process. Recognizing alterations in these factors is an important early clue of disease recognition. While frequently it is nonspecific, in many instances it can lead directly to a diagnosis. Careful observation of the respiratory rate and pattern is a crucial part of the physical examination.Simple inspection of the respiratory cycle, observing rate, rhythm, inspiratory volume, and effort of breathing, is all that is necessary. The rate is noted by observing the frequency of the inspiratory phase, since this phase is active and easy to count. Record the number of breaths per minute; this is the respiratory rate. While observing the rate, note the inspiratory expansion of the chest cage. This expansion should be the same during each cycle.
Normally, the accessory muscles of inspiration and expiration are not used. Their use should be observed and, if found, recorded as "use of accessory muscles on inspiration" and "expiration is active with abdominal muscle contraction."
Basic Science
The respiratory system's major functions are to provide an adequate oxygen (O2) supply to meet the energy production requirements of the body and maintain a suitable acid-base status by removing carbon dioxide (CO2) from the body. This is accomplished by moving varying volumes of air into and out of the lungs. Ventilation, the process of air movement into the lungs, is a carefully controlled modality with a wide range of response that enables the markers of gas exchange adequacy (Pao2, Paco2, and pH) to be kept within a relatively small physiologic range.To maintain accurate control the respiratory system has a central respiratory pacemaker located within the medulla of the brainstem. Neural output travels from this center through the spinal cord to the muscles of respiration. The changes are effected through two groups of muscles, inspiratory and expiratory, which contract and relax to produce a rhythmic respiratory rate and pattern. In most individuals with unchanging metabolic demand, the rate and pattern are surprisingly constant, only interrupted every several minutes by a larger inspiratory effort or sigh. Ventilation at rest in most individuals requires only the inspiratory muscles. Expiration is usually passive and is secondary to the respiratory system returning to its resting state. Therefore, with quiet breathing the inspiratory time is the period of active respiratory pacemaker output. Adjusting the rate, length, and intensity of neural output from the pacemaker will lead to changes in the breaths per minute and the volume of each inspiration or tidal volume. These final outputs of the respiratory pacemaker, the rate and tidal volume, are the two components of ventilation. The expiratory muscles begin to play a role with disease or increased ventilatory demands. When this occurs, the length of time it takes to empty the lungs adequately will also lead to changes in rate and tidal volume.
Minute ventilation is the product of rate and tidal volume. It is important to differentiate between the effect changes in rate and tidal volume have on gas exchange. Any given tidal volume is divided into two components. One part is the dead space. This is the portion of the volume moved into the lungs during ventilation that does not come into contact with functioning pulmonary capillaries. An example is air at the end of inspiration, which reaches only the trachea or bronchi where there are no capillaries. Since there is no air–blood interface, O2 cannot reach the circulation nor can any CO2 be removed. The other component is called the alveolar volume. This is the part of a tidal breath that enters the air spaces of the lung that are perfused by functioning capillaries. In normal individuals these air spaces are the terminal respiratory unit and include the respiratory bronchioles, alveolar ducts, alveolar sacs, and alveoli. Only the alveolar volume component of each tidal breath contributes to gas exchange; the rest is really wasted ventilation. If minute ventilation is increased by making the tidal volume larger, it will have a greater effect on gas exchange than if the same minute ventilation is reached by increasing the rate.
Regulation of Pacemaker Output
The medullary respiratory control center, or pacemaker, receives three kinds of feedback. These impulses are integrated within the control center. The output from the respiratory center is then altered in timing or intensity, leading to changes in the rate and tidal volume. The three kinds of feedback are chemical, mechanical, and input from higher cortical centers.The normal individual is able to keep Pao2, Paco2, and pH within narrow limits. In order to accomplish this level of control, the respiratory center receives input from both peripheral and central chemoreceptors. The major peripheral receptors are located within the carotid bodies found in the bifurcation of each common carotid artery. There are also similar structures in the aorta, but less is known about these aortic bodies. The afferent limb of these receptors responds to Pao2 and pH changes. The efferent limb produces changes in minute ventilation through the respiratory control center. The response to changing Pao2 levels can be detected as high as 550 mm Hg, but at that Pao2 level the resulting change in rate and volume is small. As the Pao2 drops to 55 to 60 mm Hg, there is a much greater and more important respiratory response with large increases in minute ventilation for each mm Hg change in Pao2. The carotid body also responds to small changes in pH, but approximately two-thirds of the response to pH is a result of the central chemoreceptors. The response of the carotid bodies to Paco2 is secondary to changes in pH resulting from the Paco2 change.
The medullary chemoreceptor is located on the ventral surface of the medulla. This receptor responds to changes in pH and is the most important receptor regarding respiratory changes to acid-base alterations. It responds to changes in cerebrospinal fluid rather than blood and is very sensitive to very small changes in the hydrogen ion concentration in the cerebrospinal fluid. Since CO2 rapidly crosses the blood-brain barrier, it rapidly alters the pH of the spinal fluid. An increase of 1 mm Hg of Pco2 in the cerebrospinal fluid leads to an increased ventilation of 2 to 3 liters per minute. The medullary chemoreceptor also adjusts the respiratory response to altered pH secondary to metabolic acidosis or alkalosis. With the slower equilibration of hydrogen or bicarbonate across the blood-brain barrier, however, these changes are not as quick as the rapid respiratory changes produced by a change in Pco2.
Another neural input to the respiratory pacemaker comes from receptors in the lung and is related to the mechanical properties of the lung. An individual who elected to breathe at a rate of 5 breaths per minute with a large tidal volume would have efficient gas exchange because the ratio of dead space to tidal volume would be low. The larger the inspiratory lung volume, however, the greater becomes the elastic recoil of the lung. At greater lung volumes, chest wall elasticity is also added and must be overcome. Therefore, the larger the inspiratory lung volume, the greater the inspiratory pressure needed to overcome the elastic recoil and expand the lung. The greater the inspiratory pressure, the greater is the work of breathing by the respiratory muscles. The respiratory system appears to choose a rate that requires the least amount of mechanical work while maintaining adequate gas exchange. There is a wide range of tidal volumes before the mechanical limitation comes into effect, but there appear to be lower limits of rates that are not tolerated because of the required increase in inspiratory work.
Receptors in the lung itself appear to contribute to this inspiratory limitation. One group is the stretch receptors. Efferents from these increase their neural output the larger a given lung volume becomes. In some animals these reflexes are very important, but in normal humans they appear to be less important and can be easily overcome by other neural inputs. The output of these receptors can, however, limit the degree of inspiration by means of the Hering-Breur reflex. Probably in disease states this reflex plays a more important role in limiting inspiration. The other lung parenchymal receptors that may play a role in limiting the size of each tidal volume are the juxtapulmonary capillary receptors, or J-receptors. These receptors fire when pulmonary capillaries are distended.
A final modulator of the central respiratory drive is input from higher centers. For example, the state of being awake is associated with important neural inputs to the respiratory center that will play a large role in determining an individual's respiratory rate and pattern. When an individual falls asleep, the cortical input decreases, as does the respiratory center output. During nondreaming or non-rapid-eye-movement sleep, the input from the chemical receptors becomes increasingly important. If absent, apnea may result. During sleep associated with rapid eye movements or dreaming, the breathing patterns may be related to the contents of the dreams and again reflect input from higher cortical centers. Higher center input also accounts for hyperventilation associated with anxiety and other behavioral factors.
Alterations in Rate and Tidal Volume
All three types of input are integrated in the medullary respiratory center and lead to changes in the minute ventilation. These changes are seen as changes in rate, volume, or both. Table 43.1 demonstrates how these factors may interrelate in normal individuals. If the ventilation during a minute (minute ventilation) was 6 liters and was done at a rate of 60 breaths with a tidal volume of 100 cc each, there would be no alveolar ventilation at all. This is because the normal dead space consisting of the trachea and some bronchi is about 150 cc. Even if the rate was slowed to 30 breaths per minute, the result would be an alveolar ventilation of only 1.50 L. This would be inadequate to meet CO2 production and would lead to an elevation of the Paco2 and a lowering of the pH. Both the central and peripheral chemoreceptors would be stimulated. There might also be a concomitant fall in Pao2, which would lead to increased neural output from the carotid bodies. The result of the increase of input to the central center would be an alteration in the rate and pattern of breathing.A third breathing alternative would be choosing a respiratory rate of 2 per minute. This would give an extremely efficient breath with very little wasted as dead space. The problem with the large volume is it would require increased work of breathing and stimulate stretch receptors. Therefore, unless a constant conscious effort was maintained, the respiratory central center would inhibit inspiratory effect before reaching tidal volumes of 3 liters. Furthermore, the very efficient gas exchange could lead to a lowered Paco2. The resulting increased pH would produce less drive to breathe and lower minute ventilation. The best alternative in a normal individual would be to choose an intermediate rate of 10 to 20 breaths per minute. The example of 15 breaths per minute would meet metabolic needs effectively.
In normal individuals, multiple factors affect the respiratory rate and pattern at rest. Normal people also must adjust to changing metabolic demands, as seen with exercise. Using the input from various receptors, the respiratory center finely adjusts both rate and pattern to keep Pao2 and pH within a relatively small range in spite of increased metabolic demands of 15 or more times the needs at rest.
Clinical Significance
Abnormal Central Respiratory Control
Altered respiratory rate and pattern often accompany a variety of disease states. These diseases frequently lead to alterations in one of the three kinds of feedback to the central respiratory control center or in the control center itself. For example, pathological conditions altering Pao2, Paco2, or pH can obviously alter the input from both the carotid body and the medullary chemoreceptors. The usual response to any altered chemoreceptor input is, first, a change in tidal volume, followed by change in respiratory rate. Therefore, lung diseases that cause acute hypoxemia to a level lower than 55 to 60 mm Hg will usually produce increased ventilation. The response to increasing Paco2, and lowered pH can produce rapid changes in minute ventilation also by stimulating the chemoreceptors. The elevation of Paco2 is not always associated with the expected increase in minute ventilation, however. Elevation of PaCO2, can lead to CO2 narcosis and depression of the respiratory center. Metabolic acidosis, in contrast, will most likely increase the ventilation predominantly by increasing the tidal volume. Kussmaul respiration, the classic pattern seen in diabetic ketoacidosis, consists of slow, deep breaths that reflect the increased tidal volume and actual slowing of rate. The occurrence of respiratory compensation for a metabolic change may be slowed because cerebrospinal fluid changes lag behind blood changes. An acute pH drop in the blood will stimulate the peripheral chemoreceptors, leading to hyperventilation and acutely lowering the Paco2. This will lead to a lowered Pco2 in the cerebrospinal fluid. Since the hydrogen ion associated with the metabolic acidosis does not cross the blood-brain barrier immediately, the medullary chemoreceptors may initially reduce respiration because the reduced Pco2 will make the cerebrospinal fluid alkalotic. In a matter of hours the spinal fluid pH is decreased, and the appropriate response will occur. Ventilation will also be slowed with metabolic alkalosis. This change may not be detectable on routine physical examination. Nevertheless, it can also lead to enough slowing that the Paco2 becomes markedly elevated.Mechanical properties also are altered by diseases. Interstitial disease probably enhances the stretch receptor response, leading to rapid shallow ventilation. This is efficient because the lung with interstitial disease is less compliant, requiring more distending pressure per unit of volume than normal. By breathing at lower tidal volumes, the work of breathing is diminished. Congestive heart failure can produce a similar effect. This is partly related to the reduced compliance, but also may be a result of stimulation of the J-receptors, which lie right next to the capillaries.
The respiratory center feedback from the higher cortical centers can also be modulated with diseases. Anxiety can increase the respiratory rate and pattern. The acute hyperventilation syndrome is an example where drive from higher centers can maintain a high minute ventilation in face of an elevated pH. Increased intracranial pressure leads to a rapid and deep breathing pattern. This pattern is frequently seen with head trauma. Pain contributes to a rapid respiratory rate. A fractured rib produces pain on inspiration and therefore leads to a low-volume, rapid-rate pattern. Tachypnea is commonly part of any chest pain and is partly modulated through higher cortical input.
The central controlling center can be affected directly. Any central nervous system depressing drug will reduce the respiratory rate and pattern. It will also blunt the response to other neural inputs. The patient with obstructive lung disease who receives a narcotic frequently will elevate the Paco2 even further. The same is true for many drug overdoses. If the central nervous system is depressed by drugs, the depression of the respiratory center leads to CO2 retention.
Central Nervous System Abnormalities
The changing rate and pattern of respiration can often suggest localization of CNS changes. Understanding of the areas of the brain involved with specific patterns have come from animal studies. Lesions or cuts made in various parts of the brain lead to specific breathing patterns. Transection of the pons will not affect normal breathing if the vagi are intact. If vagi are cut, however, larger tidal volumes with a slower rate are observed. A midpons transection will lead to maintenance of spontaneous breathing but with a slow and regular pattern. If the vagi are cut, apneustic breathing occurs. This is sustained inspiratory spasm. Pontomedullary junction transection will lead to an irregular, ataxic breathing pattern.Abnormal Respiratory Patterns
Cheyne-Stokes breathing is a classic breathing pattern seen in both normal individuals at altitude and individuals with severe neurological or cardiac disease. The pattern (Figure 43.1) demonstrates periods of hyperventilation alternating with periods of apnea. The apneic spells can last as long as 45 seconds. The abnormality appears to be related to a slow feedback loop and an enhanced response to Paco2. During periods of hyperventilation, the Paco2 is at its highest, while the Pao2 is at its lowest. As ventilation slows, the Paco2 drops and reaches its lowest level during apnea. It is important to observe that Paco2 levels do not exceed the normal range during any part of the cycle.There are several probable causes of this abnormal breathing pattern. Many cases have diffuse cerebral damage, whereas some individuals are in congestive heart failure. It is seen in normal individuals during sleep at altitude. It has been shown in dogs that a markedly prolonged circulation time from the left ventricle back to the brain can also induce Cheyne-Stokes respiration. In individuals with either neurological or cardiac disease it frequently is a poor prognostic sign. Treatment is usually improvement of the underlying disease, but aminophylline has been effective in some cases.
A Cheyne-Stokes respiratory pattern can also be seen in individuals with much more severe neurological depression. These individuals have a low pontine or upper medullary lesion. Unlike the more classic Cheyne-Stokes respiration, these individuals are cyanotic and have CO2 retention. They have reduced sensitivity to CO2. Oxygen will enhance this pattern, while it may reduce the more classic picture.
Biot respiration, or cluster breathing, is also periodic in nature but does not have the crescendo–decrescendo pattern seen with Cheyne–Stokes respiration. It is clusters of irregular breaths that alternate with periods of apnea. This breathing pattern is seen in individuals with pontine lesions. Ataxic breathing is one of varying tidal volumes and rates. These individuals can frequently keep their rate more rhythmic if they try consciously. The abnormality is in the medullary chemoreceptor or the medullary respiratory control center.
One other aspect of respiratory pattern must be considered. This is the coordination between the chest wall and abdomen. Normal individuals contract both the diaphragm and external intercostal muscles during inspiration. On physical examination the action of both inspiratory muscle actions can be determined. The diaphragm, when contracting normally, moves the abdominal contents downward and outward. In Figure 43.2 this is represented by an upward deflection of the abdominal curve. On physical examination it is felt as an anterior movement of the abdomen. The other major groups of inspiratory muscles, the external intercostals, move the chest wall outward. This can also be determined by feeling an anterior movement of the chest wall during inspiration and is reflected in the figure as an upward deflection. In normal persons, therefore, there is a coordinated movement of the chest wall and abdomen moving outward on inspiration and inward on expiration. Alterations in this pattern will allow diagnosis of changing respiratory muscle contribution to the tidal breath.
Paralysis of the intercostals results from cervical spinal cord injury. In this group of patients on physical exam there is a paradoxical movement of the chest wall inward and the abdomen outward during inspiration (Figure 43.2). This reflects the passive movement of the chest wall. The pattern results from the tidal breath being produced by diaphragmatic contraction. Diaphragmatic paralysis can be diagnosed or suggested by an inward movement of the diaphragm during inspiration. This movement is accentuated in the supine position. In that position, diaphragmatic contraction produces approximately two-thirds of the inspiratory volume as compared to one-third in the upright position.
Diaphragmatic dysfunction can also be diagnosed with the finding of the same paradoxical inward abdominal movement during inspiration. This pattern of respiration is seen in some individuals with severe emphysema and air trapping. The air trapping leads to a low, flat diaphragm. The diaphragm no longer can contract effectively. This inability to contract is demonstrated by the inward inspiratory movement of the abdomen. The movement results from the ineffective diaphragm being pulled into the thorax during inspiration. This breathing pattern has been reported to have a predictive value for impending respiratory failure.
Another value of determining changes in chest wall abdominal breathing patterns has been seen in individuals being weaned from mechanical ventilation. They develop a respiratory alternans pattern. A series of tidal breaths alternates between a short period of abdominal inward movement during inspiration followed by a period of chest wall inward movement during inspiration. This has been demonstrated to be associated with fatigue and indicates weaning should be stopped.
Other Applications of Respiratory Rate and Pattern
Understanding respiratory rate and pattern is a very important addition in dealing with many respiratory, cardiac, and neurological diseases. Nevertheless, understanding the role of rate and tidal volume is also essential in managing individuals requiring mechanical ventilation. This is one clinical situation where the physician can directly manipulate the respiratory rate and pattern to produce the appropriate arterial blood gases. The sum of each alveolar volume over one minute is the alveolar ventilation and is inversely related to the Paco2. Hyperventilation reduces Paco2, leading to decreased respiratory drive and a patient who does not "trigger" the ventilator. Hypoventilation produces CO2 retention and increased respiratory drive by the patient. This leads to either rapid ventilation rates triggered by the patient or a patient demonstrating marked discomfort. Consideration of dead space is important. Diseases can increase the dead space by reducing the capillary bed. Therefore, larger tidal volumes may be necessary to get adequate gas exchange. Finally, volumes that are too large lead to asychrony by the patient who tries to start the next breath before the ventilator is ready. This may relate to stretch receptor responses, but is a fatiguing and inefficient mode of gas exchange.References
- Brown HW, Plum F. The neurological basis of Cheyne-Stokes respiration. Am J Med. 1961;30:849–86.
- Cohen CA, Zagelbaum G, Gross D, Roussos CH, Macklen PT. Clinical manifestation of inspiratory muscle fatigue. Am J Med. 1982;73:308–16. [PubMed: 6812417]
- Kryger MH. Abnormal control of breathing. In: Kryger MH, ed. Pathophysiology of respiration. New York: Wiley, 1981;103–22.
- Mead J. Control of respiratory frequency. J Appl Physiol. 1960;12:882–87. [PubMed: 13934289]
- Tobin MJ, Snyder JV. Cheyne-Stokes respiration revisited: controversies and implications. Crit Care Med. 1984;12:882–87. [PubMed: 6435956]
Figures
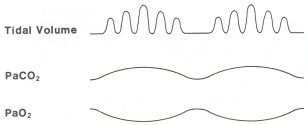
Figure 43.1
Cheyne-Stokes breathing.
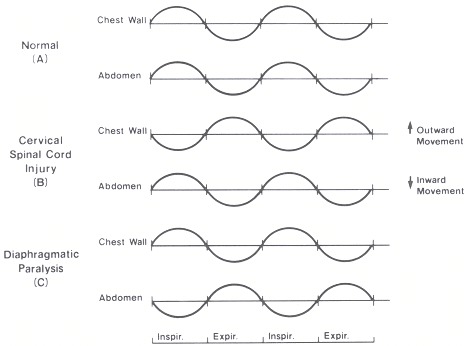
Figure 43.2
Chest wall and abdominal coordination during tidal breathing.
Tables
Table 43.1The Effect of Changes of Respiratory Rate and Tidal Volume on Alveolar Ventilation
| Minute ventilation (L) | Respiratory rate | Tidal volume (L) | Dead spacea (L/min) | Alveolar ventilation (L/min) |
|---|---|---|---|---|
| 6.0 | 60 | 0.1 | 6.0 | 0 |
| 6.0 | 30 | 0.2 | 4.5 | 1.5 |
| 6.0 | 15 | 0.4 | 2.25 | 3.75 |
| 6.0 | 2 | 3.0 | 0.3 | 5.7 |
- a
- A dead space of 150 ml/breath is assumed.
Copyright © 1990, Butterworth Publishers, a division of Reed Publishing.
Valuable Respiratory Rate is simply the number of human breaths an individual takes per minute.
Trả lờiXóa